It has always been know that the first hunter-gatherers of Europe were SSAs (Sub-Saharan Africans) . Now we can declare that the carriers of haplogroup R1, that introduced the European agro-pastoral cultures to Europe, i.e., Bell Beaker and Yamnaya were probably also SSAs because they carried V88.
This should not be surprising according to Turek, the Bell Beaker culture probably began in Morocco. Neolithic migrants into Europe from the Levant were also SSA. Trenton W. Holliday, tested the
hypothesis that if modern Africans had dispersed into the Levant
from Africa, "tropically adapted
hominids" would be represented in the archaeological history of the Levant, especially in relation to the Qafzeh-Skhul hominids. This researcher found that the Qafzeh-Skhul
hominids (20,000-10,000),were assigned to the Sub-Saharan population, along
with the Natufians samples (4000 BP).
Holliday also found African fauna in the area. If they were Sub-Saharan Africans in the Levant.
Toomas Kivisild1 (2017).The study of human Y chromosome variation through ancient DNA.
web page provides a detailed discussion of R1 in prehistoric Europe.
The article is interesting. It is most interesting because it places V88 in ancient Europe. It is sad that researchers fail to publish this reality.
Kivisild (2017) also made it clear that V88 is the earliest offshoot of R-M343 .
quote:
Late Neolithic, Early Bronze Age and Iron Age samples from Central and Western Europe have typically the R1b-L11, R1a1-Z283 and R1a-M417 (xZ645)
affiliation while the samples from the Yamnaya and Samara neighbourhood are different and belong to sub-clades R1b11-Z2105 and R1a2-Z93 (Allentoft et al. 2015; Cassidy et al. 2016; Haak et al. 2015; Mathieson et al. 2015; Schiffels et al. 2016).

The R1b11-Z2015 lineage is today common in the Caucasus and Volga-Uralic region while being virtually absent in Central and Western Europe (Broushaki et al.2016). Interestingly, the earliest offshoot of extant haplogroup R1b-M343 variation, the V88 subclade, which is currently most common in Fulani speaking populations in Africa (Cruciani et al. 2010) has distant relatives in Early Neolithic samples from across wide geographic area from Iberia, Germany to Samara (Fig. 7).
The presence of the carriers of V88 in Europe makes it clear that Sub-Saharan Africans had been in Europe for an extended period of time. Moreover it is clear that 25kya SSAs carrying haplogroup R1 were in Eurasia, Africa and the Americas.
.

The Basal Eurasians that are mixed into modern Europeans, who came from the Middle East, and North Africa were SSA's not Indo-European speakers.
.
quote:
Interestingly, the earliest offshoot of extant haplogroup R1b-M343 variation, the V88 sub-clade, which is currently most common in Fulani speaking populations in Africa (Cruciani et al. 2010) has distant relatives in Early Neolithic samples from across wide geographic area from Iberia, Germany to Samara (Fig. 7).
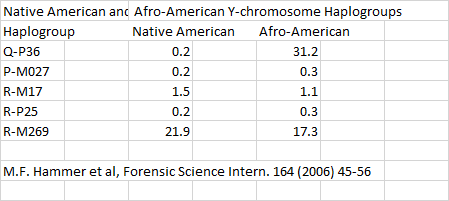
This quote makes it clear the V88 sub-clade, had relatives in Early Neolithic samples from across wide geographic area from Iberia, Germany to Samara. This would place carriers of V88 among the Yamnaya and Bell Beaker people. Given the wide distribution of M269 in Africa, the carriers of this haplogroup were probably also Africans since the Bell Beaker people/culture originated in Morocco as noted by Turek (2012) and the Neolithic people of the Levant were also SSA as proven by Holliday (2000).
Reference:
Holiday, T.
(2000). Evolution at the Crossroads: Modern Human Emergence in Western Asia, American Anthropologist,102(1) .